|
Gaining
and Losing Biodiversity
Island
biogeograpy | Measuring diversity | Gaining
biodiversity | Losing biodiversity
Globally, diversity naturally
has increased over time, though the great mass extinctions have decreased
it for a while. The most famous of the mass extinctions is the one that
claimed the dinosaurs, but we are currently in the midst of a human-created
mass extinction. Local diversity, on the other hand, is constantly increasing
and decreasing at very short time scales. There are many factors that
affect diversity, and the major natural circumstances are given here.
Human-generated impacts on diversity have almost always been negative,
and are covered in the Conservation Issues section.
Island
Biogeography
One of the first major theories
of biodiversity, the theory of island biogeography, formulated by R. MacArthur
and E.O. Wilson applies to the patterns of diversity found on islands.
In it, islands start out empty of species, who arrive from a large area
(referred to as the mainland, though it doesn't have to actually be a
continent) and from neighbouring islands.
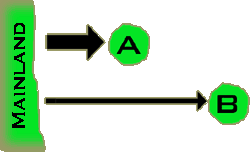 The
chance that a species will land on the island depends mostly on the distance
that the island is from the mainland; the greater the distance the less
often a species will find its way to the island. In the figure to the
right, island A is closer to the mainland than island B, and more species
find their way to it. Species on smaller islands have smaller populations,
making them more vulnerable to extinction. The number of species present
on the island is a balance between the rate at which new species arrive
and old species go extinct on the island. The
chance that a species will land on the island depends mostly on the distance
that the island is from the mainland; the greater the distance the less
often a species will find its way to the island. In the figure to the
right, island A is closer to the mainland than island B, and more species
find their way to it. Species on smaller islands have smaller populations,
making them more vulnerable to extinction. The number of species present
on the island is a balance between the rate at which new species arrive
and old species go extinct on the island.
This theory is neutral, meaning
that all species are considered to be equal. In reality, some species
are better at dispersing than others and are thus more likely to be found
on islands. The exact species that are actually present has been found
to be fairly random, though, and the theory does a good job of predicting
the number of species to be found. What the theory calls islands doesn't
have to be actual islands; lakes are effectively islands, as are isolated
patches of habitat, and the theory has been extended to deal with peninsulas,
bays, and other only partially isolated areas.
Measuring
Diversity
To detect changes in biodiversity
there has to be a way to measure it. Although at first glance biological
diversity seems to be an obvious idea, quantifying it is much more difficult.
Making an attempt to express it as a single number is futile, as a single
number cannot hope to convey the different components. There are three
common ways to measure diversity:
Numbers:
It
is possible to measure how many species are found in an area,
or how many alleles
(defined above) a species
has for a single locus, or how many functional groups (defined below)
or taxonomic groups
higher than species are present in an ecosystem. This is considered
a reasonable if incomplete way of measuring diversity, and can be expressed
as the number of species found per unit area, per unit mass, or per
number of individuals identified. What controversy exists about this
component is mainly about how to standardize measures that are taken
at different scales.
Evenness:
If almost
every individual in
an area is from the
same species, the diversity would not seem high, even if there are many
species present. Evenness
measures to what extent individuals are evenly distributed among species
(if one is looking at the species level). The most common values that
are used are species number and species
evenness. How to represent even these two components as a single number
has been controversial (see Magurran 1988 and Smith and Wilson 1996
for examples of indices and the controversy), and the number of different
abundance indices is large, although certain ones (the Shannon and Simpson
indices, for instance) are far more commonly used than others.
Difference:
A site with many species
is considered to have high diversity, but what if those species are
all very closely related? If another site had fewer species, but those
species were more distantly related, would that second site have a lower
or higher diversity? Measuring the evolutionary distance between the
different units is important, as it is on a different level than something
like species number, which doesn't measure how different the species
are. Measurements of difference include disparity and character diversity.
Three sample areas are given
to the above right, each of which is most diverse in a different way.
The top area (area 1) has the greatest number of species, four in total.
But half of the individuals in the sample are from the same species. The
middle area (area 2) has fewer species, only three, but it has a greater
evenness; there is an equal chance of getting an individual from each
of the three species. The bottom area (area 3) has even fewer species,
just two, but it has the greatest difference. While the other samples
contain only insect species, this one contains both insects and a mammal,
which is very distantly related to insects.
Gaining
Biodiversity
Mutation
Mutations increase genetic diversity by altering the genetic material
(almost always DNA) of organisms. Once mutations arise, they are passed
on to the mutated organism's offspring, and in time may either disappear
if the line dies out. Depending upon the specific mutation, the result
can range from no effect whatsoever to the creation of an entirely new
species. Although this gives rise to differences in organisms, it is an
extremely slow process compared to the other ways in which local diversity
increases. Ultimately, though, this is the only way in which diversity
is truly created.
Speciation
The creation of a new species is known as speciation. Species are typically
defined as being unable to successfully breed with other species (the
so-called Biological Species Concept), although there are other ways of
defining species. The origin of new species naturally has the largest
immediate effect on species-level diversity; the immediate changes to
genetic and ecosystem diversity are usually minimal, though the effects
will grow in time. Speciation can occur through several different means,
including geographical isolation, competition, and polyploidy. These are
described below.
Geographical Isolation:
Geographical isolation, such as new mountain chains or a lake whose
level lowers enough that it splits into two separate lakes, can divide
a population into two separate populations. The two isolated populations
continue to evolve separately from one another. Eventually they can
diverge to a great enough degree, either through adaptation to their
differing environments or through random mutations, that they are no
longer able to interbreed and are considered to be different species.
Competition: If a
new resource, such as a new food source, becomes available to a population,
some part of the population may become specialized in obtaining that
resource. Being specialized in obtaining either the new resource or
the original resource may be better than trying to obtain both. If so,
then the specialists would be better off mating with the other specialists
on the same resource, as mating with someone who uses the other resources
will result in offspring that aren't specialized for either resource
and at a disadvantage. In time, there is a chance that the population
will split into two species, each specialized on one of the two resources.
This can happen, but it is probably a fairly rare event.
Polyploidy: Speciation
through polyploidy happens far more often in plants than in animals,
as animals are much more sensitive to large changes in their genetic
structure. Most species are diploid, meaning they have two ("di" meaning
two) copies of each chromosome (large packages of DNA), one from each
of their parents. An individual in a normally diploid species may have
more copies of these chromosomes, being polyploid ("poly" meaning many),
through errors at the cellular level. The additional copies of the chromosomes
render them unable to produce functional offspring with normal members
of their species. Plants often fertilize themselves to at least some
extent, so polyploid species can arise from a single individual. This
method of speciation is almost instantaneous, happening in a single
generation, and is more common in plants than animals.
Immigration
 Immigration
increases diversity as new individuals and perhaps even new species enter
an area, increasing its diversity. The rate at which immigration happens
depends on the size of the area in question, how many species are there
already, and how close the area in question is to the source of immigration.
Even if a species is unable to survive in an area, a constant flow of
immigrants to the area can keep the species present indefinitely. Island
biogeography is the classic theory on the topic of how these factors affect
immigration and more, and is explained above. Immigration
increases diversity as new individuals and perhaps even new species enter
an area, increasing its diversity. The rate at which immigration happens
depends on the size of the area in question, how many species are there
already, and how close the area in question is to the source of immigration.
Even if a species is unable to survive in an area, a constant flow of
immigrants to the area can keep the species present indefinitely. Island
biogeography is the classic theory on the topic of how these factors affect
immigration and more, and is explained above.
Most species that immigrate
to a new ecosystem have only minor effects on their new system, though
some drastically change it. Zebra mussels, native to the Caspian Sea and
Ural river, were first recognized in the Great Lakes in 1988. It is most
likely that they were brought over in ballast water. Since then they have
spread throughout the Great Lakes and beyond, killing native mussel populations
and fouling all manner of pipes and intakes.
Succession
Succession is the process through which an area gains species as successive
communities of organisms replace one another until an endpoint is reached.
This endpoint, or climax community, is commonly a forest in southern Canada.
Succession may begin on bare rock, an abandoned field, the burned remnants
of a forest, or any stage before the endpoint. A hypothetical bare field
isn't bare for long before annual plants appear. They are replaced within
a few years by perennial plants and shrubs, who in turn are replaced by
pine trees. Eventually, hardwood trees invade and replace the pines, forming
the hardwood climax community.
Different regions have varying
climax communities; the tundra of the north is extremely different from
the grasslands of the prairies or the west coast rainforests, though they
are all the local endpoints of succession. One usually refers to the different
stages of succession in terms of the plants rather than the animals because
the plants precede the animals and provide the structure and environment
that the animals live in. One exception to this is aquatic communities,
where sponges, corals, bivalves and other animals are responsible for
much of the three-dimensional structure of the community. The climax community
is typically the most diverse stage of succession, and each stage of succession
is more diverse than the one preceding it. This pattern depends on the
group being looked at; plant diversity actually decreases at the final
stage, while animal diversity increases to the end. Species that were
common in the early stages of succession will not be common in the later
stages, but may still be found if small disturbances in the area effectively
set the disturbed area back to an earlier successional stage (see the
page on Abundance and Composition for more details).
Losing
Diversity
Extinction
 Extinction
is more an outcome than a process. Once a species goes extinct, all the
diversity that it represented is lost forever. The vast majority of species
that have ever existed are now extinct through natural processes, whether
by mass extinction or by the more common individual extinction. Genes
also go extinct if they fail to get passed on to the next generation,
though it's not necessary for the entire species to go extinct as well.
Ecosystems may be destroyed by severe disturbances, but they don't really
go extinct unless the species that make them up are lost. Extinction
is more an outcome than a process. Once a species goes extinct, all the
diversity that it represented is lost forever. The vast majority of species
that have ever existed are now extinct through natural processes, whether
by mass extinction or by the more common individual extinction. Genes
also go extinct if they fail to get passed on to the next generation,
though it's not necessary for the entire species to go extinct as well.
Ecosystems may be destroyed by severe disturbances, but they don't really
go extinct unless the species that make them up are lost.
Species can also go locally
extinct; in this case, they are said to be extirpated. Although the local
loss of diversity is the same, the species still exists elsewhere and
may be able to return in the future through immigration. Much the same
thing can happen to genetic diversity, as particular alleles are lost
in a population.
Competition
If one species outcompetes others to a dramatic extent, the result may
be extirpation or perhaps even extinction of the other species and a reduction
of diversity. Diversity, in the sense of evenness, will also be lowered
if other species have their populations greatly reduced by a competitor
or predator, even if the species aren't extirpated. As species that have
been eliminated simply aren't around, it's rare to see this process happening
unless a species has recently invaded or conditions have recently changed.
Disturbances
Disturbances can maintain diversity (see Abundance and Composition, below),
but extremes can reduce diversity. Constant large-scale disturbance can
eliminate many populations and keeps an area at the early levels of succession,
which have lower diversity (see above). An area with no disturbances at
all would end up completely at the final stage of succession. This would
prevent the presence of the species that would normally be found at intermediate
stages of succession, living in the disturbed areas.
Bottlenecks
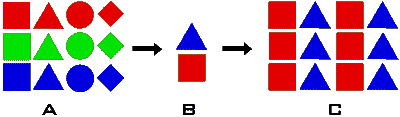 Genetic
bottlenecks happen when many individuals in a population die. In the example
to the right, the population initially has many different types of shapes
and colours, representing genetic diversity (A). The few individuals that
are left after most die (B) have a small amount of the genetic diversity
that originally existed, as much of the genetic diversity was lost with
the rest of the population. Although the population's numbers quickly
recover (C), the genetic diversity is much slower to respond, which can
cause problems if conditions change in the future, as the reserves of
diversity that would be useful won't be there. Genetic
bottlenecks happen when many individuals in a population die. In the example
to the right, the population initially has many different types of shapes
and colours, representing genetic diversity (A). The few individuals that
are left after most die (B) have a small amount of the genetic diversity
that originally existed, as much of the genetic diversity was lost with
the rest of the population. Although the population's numbers quickly
recover (C), the genetic diversity is much slower to respond, which can
cause problems if conditions change in the future, as the reserves of
diversity that would be useful won't be there.


|
|






