|
Enrichissement
et appauvrissement de la diversité
La
biogéographie insulaire | La mesure de la diversité
| L'enrichissement de la diversité | L'appauvrissement
de la diversité
Au niveau planétaire, les processus
naturels induisent spontanément une accentuation de la diversité au fil
du temps, et ce, même si les extinctions massives peuvent la faire temporairement
régresser. La plus célèbre de ces extinctions massives est bien sûr celle
qui a rayé les dinosaures de la surface du globe. N'oublions pas pour
autant que nous nous trouvons actuellement au plein cur d'une autre extinction
massive causée, celle-là, par l'homme. Au plan local, par contre, la diversité
augmente et décroît constamment à brève échéance. Plusieurs facteurs influent
sur la diversité. Nous rappelons ci-dessous les circonstances naturelles
les plus déterminantes. En ce qui concerne les impacts exercés par l'homme
sur la diversité, jusqu'à présent, ils se sont presque tous révélés négatifs.
(Pour en savoir plus à ce sujet, voir la section sur la conservation.)
La
biogéographie insulaire
La théorie de la biogéographie
insulaire a été l'une des premières théories majeures de la biodiversité.
Énoncée par R. MacArthur et E. O. Wilson, elle s'applique aux schèmes
de la diversité que l'on observe sur les îles. Elle stipule que les milieux
insulaires sont à l'origine vides d'espèces et que celles-ci y arrivent
peu à peu en provenance de régions vastes (désignées sous le terme de
« continents », bien qu'il ne s'agisse pas forcément de continents à proprement
parler) ou d'îles voisines.
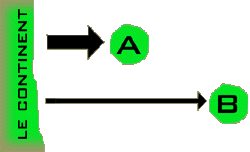 La
probabilité qu'une espèce atterrisse sur une île est fonction essentiellement
de la distance entre cette île et le continent. Plus cette distance est
grande, moins les espèces étrangères ont de chances de « découvrir » l'île
et de s'y implanter. Par ailleurs, les espèces établies sur de petites
îles constituent des populations restreintes, ce qui les rend plus susceptibles
d'extinction. Sur une île donnée, le nombre des espèces présentes dépend
de la fréquence à laquelle les nouvelles espèces arrivent et de la fréquence
à laquelle les anciennes disparaissent. La
probabilité qu'une espèce atterrisse sur une île est fonction essentiellement
de la distance entre cette île et le continent. Plus cette distance est
grande, moins les espèces étrangères ont de chances de « découvrir » l'île
et de s'y implanter. Par ailleurs, les espèces établies sur de petites
îles constituent des populations restreintes, ce qui les rend plus susceptibles
d'extinction. Sur une île donnée, le nombre des espèces présentes dépend
de la fréquence à laquelle les nouvelles espèces arrivent et de la fréquence
à laquelle les anciennes disparaissent.
Cette théorie est neutre en
ceci qu'elle considère toutes les espèces comme égales. En réalité, certaines
espèces sont mieux outillées que d'autres pour conquérir de nouveaux territoires,
donc elles sont plus susceptibles de coloniser les milieux insulaires.
Les recherches sur le terrain montrent cependant que l'agencement des
espèces effectivement présentes sur les îles semble s'être défini de manière
passablement aléatoire, et la théorie prédit avec une exactitude raisonnable
le nombre d'espèces présentes. Ce que cette approche désigne sous le vocable
d'« île » n'est pas nécessairement une île au sens propre du terme. Les
lacs peuvent être assimilés à des milieux insulaires, de même que des
fragments d'habitat isolés. La théorie de la biogéographie insulaire a
d'ailleurs été étendue aux péninsules, aux baies et à d'autres régions
qui ne sont isolées que partiellement.
La
mesure de la diversité
Pour détecter les changements
qui touchent la biodiversité, il faut pouvoir la mesurer. À première vue,
la diversité biologique semble être une notion évidente, facile à appréhender.
Quand on y regarde de plus près, toutefois, on constate qu'il n'est pas
si aisé de la quantifier. Tenter de l'exprimer par un chiffre unique serait
chose vaine : une seule grandeur ne peut pas rendre compte de toutes ses
composantes. Dans les faits, on dispose essentiellement de trois méthodes
pour mesurer la diversité.
Les indicateurs
chiffrés:
On peut mesurer le nombre
des espèces présentes dans une région, ou le nombre d'allèles (voir
ci-dessus) qu'une espèce possède pour un même locus (un même emplacement)
ou encore, le nombre des groupes fonctionnels (voir ci-dessous) ou des
groupes taxonomiques d'un rang supérieur à celui de l'espèce qui sont
présents dans l'écosystème. Considérée généralement comme valable, quoique
incomplète, cette méthode quantitative permet de mesurer la diversité
au nombre des espèces par unité de superficie ou pour l'ensemble des
individus identifiés. Les doutes qu'elle suscite concernent essentiellement
la difficulté de ramener sur un même plan des mesures prises à différentes
échelles.
L'équitabilité:
Si presque tous les individus
d'une région appartiennent à une même espèce, la diversité est a priori
faible, et ce, même si la région compte par ailleurs un nombre élevé
d'espèces. L'équitabilité désigne l'égalité plus ou moins grande de
la répartition des individus entre les différentes espèces (si l'on
se place au niveau spécifique). Les grandeurs les plus utilisées dans
ce domaine sont donc le nombre des espèces mais surtout, leur homogénéité
numérique. Cependant, les experts ne s'entendent pas sur la méthode
à adopter pour exprimer ces deux grandeurs au moyen d'un seul chiffre
(pour en savoir plus sur les indices et sur la controverse entourant
cette méthode, voir Magurran, 1988, et Smith & Wilson, 1996). Nombreux
sont donc les indices d'abondance qui sont utilisés dans le domaine,
mais certains (ceux de Shannon et de Simpson, par exemple) sont quand
même beaucoup plus couramment employés que les autres.
L'hétérogénéité:
Les régions qui abritent
de nombreuses espèces sont réputées receler une grande diversité. Mais
est-ce vraiment le cas si toutes ces espèces sont apparentées ou très
proches les unes des autres? Si une autre région regroupe moins d'espèces
mais que celles-ci sont plus disparates, son degré de biodiversité est-il
plus ou moins élevé que celui d'une région plus homogène? La mesure
de l'écart qui sépare les différentes unités du point de vue de l'évolution
est cruciale en ceci qu'elle se place sur un autre plan que le simple
comptage des espèces, par exemple, lequel ne permet pas de définir le
degré d'hétérogénéité entre les espèces présentes. Les mesures d'écarts
portent par exemple sur la disparité et sur la diversité des caractères.
On voit à droite la représentation
de trois régions. Chacune d'elles est la plus diversifiée des trois, selon
le point de vue auquel on se place. La région du haut (région 1) est celle
qui possède le plus grand nombre d'espèces (quatre), mais la moitié des
individus de l'échantillon appartiennent à la même. La région du milieu
(région 2) est celle qui abrite le moins d'espèces (seulement trois),
mais c'est aussi celle qui présente la plus grande équitabilité, puisque
les trois espèces sont également représentées. La région du bas (région
3) possède encore moins d'espèces (seulement deux), mais elles sont plus
éloignées l'une de l'autre que celles des régions précédentes. Alors que
les deux autres hébergent uniquement des insectes, celle-ci regroupe des
insectes et un mammifère, une espèce très différente des insectes.
L'enrichissement
de la diversité
Les mutations
Les mutations modifient le matériel génétique (presque toujours l'ADN)
des organismes, renforçant ainsi la diversité génétique. Quand une mutation
se produit dans un organisme, celui-ci la lègue à ses descendants, and
in time may either disappear if the line. Le résultat est variable selon
la mutation considérée. Dans certains cas, l'impact est nul; dans d'autres,
il débouche sur la création d'une toute nouvelle espèce. Ce processus
de différenciation des organismes est extrêmement lent comparé aux autres
modes d'accroissement de la diversité locale. À terme, cependant, les
mutations constituent la seule véritable source de nouveauté du point
de vue de la diversité.
La spéciation
La spéciation est la création d'une nouvelle espèce. L'espèce se définit
généralement comme une famille d'organismes capables de se reproduire
entre eux mais incapables de se reproduire avec des représentants d'autres
espèces. (C'est ce que l'on appelle la notion d'espèce biologique.) Cette
définition, toutefois, n'est pas la seule qui existe. Les experts utilisent
aussi d'autres critères pour circonscrire la notion d'espèce. Quand une
nouvelle espèce voit le jour, c'est bien évidemment sur la diversité spécifique
qu'elle exerce l'impact le plus fort à court terme. Les impacts sur la
diversité génétique et écosystémique sont plus modestes dans l'immédiat,
mais ils tendent à s'accentuer avec le temps. Plusieurs facteurs peuvent
provoquer une spéciation, notamment l'isolement géographique, la concurrence
et la polyploïdisation, que nous décrivons ci-dessous.
L'isolement géographique
-- L'isolement géographique se produit par exemple quand une nouvelle
chaîne montagneuse émerge ou quand le niveau d'un lac baisse au point
que celui-ci se scinde en deux plans d'eau distincts. Cet événement
peut séparer une population en deux groupes, chacun d'eux continuant
d'évoluer indépendamment de l'autre. À terme, par le jeu de l'adaptation
à des conditions de vie différentes ou par mutations aléatoires, ces
deux groupes peuvent diverger à un point tel que leurs représentants
respectifs ne peuvent plus se reproduire entre eux et qu'ils doivent
alors être considérés comme formant deux espèces distinctes.
La compétition --
Quand une nouvelle ressource, par exemple alimentaire, s'offre à une
population, une partie de celle-ci peut développer une spécialisation
qui la rendra la plus apte à se procurer la ressource convoitée. En
effet, il vaut parfois mieux se spécialiser dans la conquête de l'une
des deux ressources (la nouvelle ou l'ancienne) plutôt que de tenter
de s'approprier les deux de front. Dans ce cas, les individus spécialisés
ont avantage à s'accoupler avec d'autres « experts » de la même ressource
qu'avec des non spécialistes, sinon leurs descendants risquent de s'avérer
aussi inefficaces face à la nouvelle ressource qu'à l'ancienne, ce qui
représenterait évidemment un handicap pour eux. À terme, la population
peut finir par se scinder en deux espèces spécialisées, l'une par rapport
à l'ancienne ressource et l'autre, par rapport à la nouvelle. Ce phénomène
est possible en théorie, quoique probablement assez rare dans la pratique.
La polyploïdisation
-- Les animaux étant beaucoup plus sensibles que les plantes aux modifications
majeures de leur structure génétique, la spéciation par polyploïdisation
est bien plus courante dans le règne végétal que dans le règne animal.
La plupart des espèces sont diploïdes : leurs représentants possèdent
deux jeux complets de chromosomes (de grands agrégats d'ADN), un qui
leur vient de leur mère et l'autre, de leur père. Toutefois, certains
individus d'espèces normalement diploïdes peuvent s'avérer polyploïdes,
c'est-à-dire posséder, du fait d'une anomalie cellulaire, plus de deux
exemplaires de leurs chromosomes. Cette polyploïdie les empêche d'avoir
des descendants féconds avec des représentants normaux de leur espèce.
Chez les plantes, comme elles se fécondent souvent elles-mêmes au moins
dans une certaine mesure, un seul individu peut donner naissance à une
espèce polyploïde. Ce mode de spéciation produit des résultats presque
instantanés, puisqu'ils surviennent en une seule génération, et il est
plus courant chez les plantes que chez les animaux.
L'immigration
 En
introduisant dans une aire des individus nouveaux, voire des espèces nouvelles,
l'immigration accroît sa diversité. Le taux d'immigration dépend de la
taille de la région, du nombre des espèces qui s'y trouvent déjà et de
la distance entre cette région de destination et la région source. Même
si une espèce ne peut pas survivre dans une aire donnée, un flot continu
d'immigrants peut néanmoins y maintenir sa présence indéfiniment. La théorie
de la biogéographie insulaire (voir ci-dessus) est le modèle le plus utilisé
pour analyser ce phénomène de l'immigration. En
introduisant dans une aire des individus nouveaux, voire des espèces nouvelles,
l'immigration accroît sa diversité. Le taux d'immigration dépend de la
taille de la région, du nombre des espèces qui s'y trouvent déjà et de
la distance entre cette région de destination et la région source. Même
si une espèce ne peut pas survivre dans une aire donnée, un flot continu
d'immigrants peut néanmoins y maintenir sa présence indéfiniment. La théorie
de la biogéographie insulaire (voir ci-dessus) est le modèle le plus utilisé
pour analyser ce phénomène de l'immigration.
Les espèces immigrantes n'exercent
en général qu'une incidence mineure sur l'écosystème d'accueil. Dans certains
cas, toutefois, leur impact est beaucoup plus marqué. Ainsi, la moule
zébrée (la dreissena polymorphe), originaire de la mer Caspienne et du
fleuve Oural, a été repérée dans les Grands Lacs en 1988. Elle y est probablement
arrivée par les eaux de lest des navires. Depuis, elle s'est répandue
dans tous les Grands Lacs et même plus loin, éliminant les populations
de moules indigènes et obstruant les prises d'eau et autres tuyaux de
toutes sortes.
Les successions écologiques
La succession est le processus par lequel de nouvelles espèces s'établissent
dans une région donnée. Les communautés d'organismes se remplacent les
unes les autres (se succèdent) jusqu'à ce qu'un équilibre soit atteint.
Ce point d'équilibre correspond à l'arrivée à maturité (ou climax) de
la communauté, qui est dite alors « communauté climacique ». (Dans le
sud du Canada, il s'agit souvent d'une forêt.) Le processus de succession
peut commencer sur de la roche à nu, dans un champ abandonné, sur les
restes calcinés d'une forêt ou à toute étape antérieure au point d'équilibre.
Un sol nu, en fait, ne le reste jamais longtemps. Très vite, des plantes
annuelles y poussent. En quelques années, des plantes et buissons vivaces
les remplacent, puis des pins. À terme, les feuillus colonisent la région,
remplacent les pins et constituent la communauté climacique.
La communauté climacique varie
considérablement d'une région à l'autre. Ainsi, la toundra du Nord, les
herbages des Prairies et les forêts pluviales de la côte ouest diffèrent
nettement les uns des autres, alors même qu'ils constituent tous le point
d'équilibre final des successions de leurs aires respectives. Les étapes
d'une succession se définissent en général en fonction des plantes, car
celles-ci précèdent les animaux et elles établissent la structure et l'environnement
dans lequel ils vivront. À cette règle, toutefois, les communautés aquatiques
font exception. Dans ce cas, en effet, ce sont les éponges, les coraux,
les bivalves et autres organismes animaux qui établissent l'essentiel
de la structure tridimensionnelle de la communauté. Du point de vue de
la diversité d'ensemble, les étapes de la succession sont généralement
de plus en plus complexes, la diversité culminant avec la communauté climacique.
Au niveau des groupes, par contre, ce schème d'évolution ne tient plus
forcément. Ainsi, la diversité végétale diminue dans les phases ultimes
mais la diversité animale s'enrichit. Les espèces qui étaient bien répandues
au début de la succession se raréfient vers la fin mais elles peuvent
subsister si de légères perturbations ramènent la région à une étape antérieure
de la succession (voir à ce sujet la section sur l'abondance et la composition,
plus bas).
L'appauvrissement
de la diversité
L'extinction
 L'extinction
constitue un résultat bien plus qu'un processus. Quand une espèce disparaît,
toute la diversité qu'elle représentait s'éteint avec elle d'une manière
définitive. La majeure partie des espèces qui ont peuplé la planète au
fil du temps ont aujourd'hui disparu du fait de processus naturels, soit
des extinctions massives, soit, le plus souvent, des extinctions spécifiques.
Les gènes aussi peuvent disparaître s'ils ne sont pas transmis à la génération
suivante, mais leur extinction n'entraîne pas nécessairement celle de
l'espèce. Des perturbations graves peuvent détruire des écosystèmes entiers,
mais ceux-ci ne disparaissent pas vraiment, sauf si les espèces qui les
composent disparaissent. L'extinction
constitue un résultat bien plus qu'un processus. Quand une espèce disparaît,
toute la diversité qu'elle représentait s'éteint avec elle d'une manière
définitive. La majeure partie des espèces qui ont peuplé la planète au
fil du temps ont aujourd'hui disparu du fait de processus naturels, soit
des extinctions massives, soit, le plus souvent, des extinctions spécifiques.
Les gènes aussi peuvent disparaître s'ils ne sont pas transmis à la génération
suivante, mais leur extinction n'entraîne pas nécessairement celle de
l'espèce. Des perturbations graves peuvent détruire des écosystèmes entiers,
mais ceux-ci ne disparaissent pas vraiment, sauf si les espèces qui les
composent disparaissent.
Les espèces peuvent s'éteindre
localement. (On dit alors qu'elles sont déracinées.) Dans ce cas, l'affaiblissement
de la diversité au niveau de la région est le même que si l'espèce s'était
entièrement éteinte. À un niveau plus global, toutefois, l'espèce survit
et peut, le cas échéant, immigrer et réinvestir la région désertée. Le
même phénomène se produit par rapport à la diversité génétique, quand
certains des allèles d'une population disparaissent.
La compétition
Une espèce qui livre une concurrence très féroce aux autres peut causer
leur extinction locale (déracinement), voire complète, et entraîner par
conséquent un appauvrissement de la diversité. Considérée sous l'angle
de l'équitabilité (l'égalité de la répartition des individus entre les
espèces), la diversité diminue aussi lorsqu'un prédateur ou un concurrent
réduit d'une manière importante la population d'une ou plusieurs autres
espèces, même s'il ne les déracine pas complètement. Les espèces qui ont
été éliminées n'étant plus présentes, par définition, il est rare d'observer
le phénomène. C'est toutefois possible dans le cas des espèces qui colonisent
une aire depuis peu ou dans les régions dont les conditions ont changé
récemment.
Les perturbations
Les perturbations limitées peuvent n'exercer qu'un impact mineur ou nul
sur la diversité (voir plus bas la rubrique « Abondance et composition
»). Les perturbations extrêmes, par contre, l'appauvrissent presque toujours.
Quant aux perturbations constantes à grande échelle, elles éliminent en
général un nombre élevé de populations et maintiennent la région à l'un
des stades primaires de la succession, lesquels se caractérisent par un
faible degré de diversité (voir plus haut). En l'absence de perturbations,
l'aire se développe jusqu'à se stabiliser au stade final d'équilibre de
la succession. À cette étape, les espèces qui sont normalement présentes
aux phases intermédiaires (ou dans une région des alentours qui aurait
été perturbée) ont disparu.
Les goulets d'étranglement
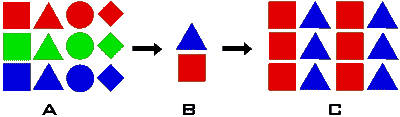 La
mort d'un nombre élevé de représentants d'une population produit ce que
l'on appelle un goulet (ou goulot) d'étranglement génétique. En d'autres
termes, ces individus emportent en disparaissant une grande partie de
la diversité génétique initiale de la population (A), ne laissant derrière
eux que quelques spécimens dépositaires d'une diversité bien inférieure
(B). L'effectif de la population peut revenir assez vite à son niveau
antérieur (C). La diversité génétique, elle, met beaucoup plus de temps
à se rétablir. Ce délai peut poser problème si les conditions de vie de
la population considérée changent, car elle ne disposera plus de la diversité
dont elle aurait besoin pour affronter ces modifications de son environnement. La
mort d'un nombre élevé de représentants d'une population produit ce que
l'on appelle un goulet (ou goulot) d'étranglement génétique. En d'autres
termes, ces individus emportent en disparaissant une grande partie de
la diversité génétique initiale de la population (A), ne laissant derrière
eux que quelques spécimens dépositaires d'une diversité bien inférieure
(B). L'effectif de la population peut revenir assez vite à son niveau
antérieur (C). La diversité génétique, elle, met beaucoup plus de temps
à se rétablir. Ce délai peut poser problème si les conditions de vie de
la population considérée changent, car elle ne disposera plus de la diversité
dont elle aurait besoin pour affronter ces modifications de son environnement.

|
|






